英国植物学家HC Watson在1835年研究植物地理分布时发现,植物种类与生境面积大小往往成正比。随后数十年,自然学家和数学家们经过进一步研究发现,物种数量与生境面积的关系遵从幂法则(S=cAz,其中S是某生境内物种数量,A是该生境面积,c和z是待估计参数)。在经历了数代自然学家的努力后,作为生态学和生物地理学中少有的经典法则之一,物种数量随生境面积空间尺度变化的幂法则关系,也就是“物种—面积关系”(SAR:Species Area Relationship)也载入了教科书中。与此同时,这一关系为研究物种灭绝以及珍惜物种保护提供了重要理论基础。显然,根据SAR理论的预测,如果自然生境面积不断减少,其中某些物种必然要灭绝。如果能够估计出某一类群(例如高等植物或哺乳动物)SAR模型的参数c和z,则有可能预测某一地区中所能容纳的高等植物或哺乳动物数量。事实上,SAR一直是保护生物学研究的重要课题之一,也是评估设置自然保护区不可或缺的最重要理论基础之一。基于物理学中时空遍历理论(Ergodic Theory),1960年代科学家提出了“物种—时间关系”(Species-Time Relationship or STR),并在古生物进化领域研究获得了一系列验证。1990年代科学家进一步扩展提出了“物种—时间空间关系” (Species-time-area relationship or STAR)。一直以来,SAR/STR/STAR 是生态学、生物地理学、保护生物学研究的热点课题之一。
SAR/STR/STAR理论有一明显局限性:忽略了物种的“丰度” (丰富程度:Abundance)。也就是说,一只大熊猫和1000只大熊猫在该理论体系中所占份量没有区别;因为该理论只考虑物种的数量(熊猫是一种动物),而不考虑具体某个物种(熊猫)的数量多少。同样,10亿只小龙虾也代表一个物种,与一只大熊猫代表的多样性没有任何区别。这一不合理的度量体系显然与人类对生物多样性重要性认识的历史局限性有关。例如,前面提到的Watson在1835年发表记载SAR专著(Watson, HC. 1835 Remarks on geographic distribution of British plants)的时间比达尔文1859年出版《物种起源》的时间早了整整24年(据维基百科:达尔文曾邀请Watson讨论进化论,Watson因不愿意旅行而谢绝了达尔文邀请,在物种起源出版后,Watson在第一时间祝贺达尔文的伟大成就)。当然,彼时人们对于生物多样性保护的必要性与今天后工业化、信息时代的认知自然无法相提并论。而生态学、生物地理学当时仅仅是自然博物学、动植物学中的一些概念。
当然,人类对生物多样性的认知一直在不断深入。科学家自然注意到了仅仅将物种数量作为生物多样性度量的严重缺陷。1948年,现代信息论创始人香农(CE Shannon)著作《通讯的数学理论》一经出版,生态学家就立即将Shannon用于度量信息的熵指标(Shannon’s Entropy) 引入生物多样性度量,而成为迄今为止最为常用的度量生物多样性的方法。Shannon于70年前创立的信息论为今天信息时代奠定了最早的理论基础,但据考证,Shannon本人并没有做过生物多样性研究,生态学家敏锐的将其成果应用于生物多样性度量。几乎是同一时间,EH Simpson于1949年在Nature发表了度量多样性的Simpson指数。1950-1980年代生态学家、数学家陆续提出了诸多多样性度量指数,使这也使得应用生态学家无所适从。前牛津大学校长、著名生态学家TRE Southwood在2000年修改再版的经典著作《生态学方法》评论到:“the result has been an ‘explosive speciation’ of diversity indices, which initially brought confusion to the subject; in addition, the ubiquitousness of some relationships and the apparent constancy of certain numerical values have added a measure of mystique.” 此后数十年中,Southwood的观点基本代表了生态学家对多样性度量方法的基本态度,那就是这些指标已近足够多了,大家似乎对这些指标优劣的争论也失去了兴趣。
此次马占山研究团队对于“生物多样性随时空尺度变化”的经典理论做出的扩展,从根本上解决了以上提到的、经典SAR/STR/STAR理论所存在的缺陷;而经典理论则成为了新理论(模型)的特例。具体可以归纳为如下五个方面:
其一、将“物种数—面积关系”(SAR)扩展到一般化的“多样性—面积关系” (DAR, Diversity-Area Relationship);采用以 Renyi 熵为基础推导出来的 “希尔数”(Hill Numbers)作为通用的“多样性指数” 取代SAR模型中的“物种数”。这一推广从根本上解决了传统SAR存在的缺陷。Renyi熵由数学家Renyi (1961)年提出,香农熵其实是Renyi 熵的特例。因此,目前常用的多样性指数:包括Shannon多样性熵指数、Simpson多样性指数、甚至“物种数(S)”(作为多样性度量指标时的正式名称“物种多度”, Species Richness), 其实都是希尔数的特例。Renyi 熵应用其实也很广泛,例如量子纠缠的信息度量、经济学中财富分布等。
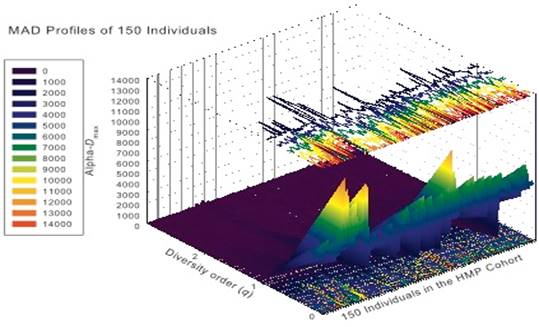
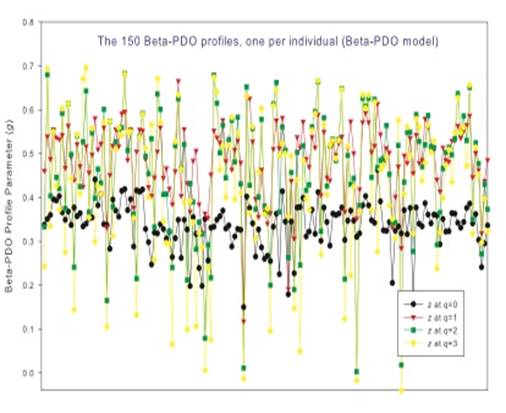
其二、“基于多样性—面积关系”(DAR),以及幂法则所具备的尺度恒定理论(Scale- Invariance) 推导出了另外两项重要参数:多样性重叠参数(PDO: Pair-wise Diversity Overlap), 以及累计多样性最大值(MAD: Maximal Accrual Diversity)。这些新推导出来的理论参数也具有重要的实际应用价值。例如,PDO可以用于估计将某一自然保护区面积扩大后多样性的增加幅度,或者保护区面积缩小后多样性的降低幅度。又如,MAD可以用于估计通过扩大保护区面积所可能取得的多样性最大理论值;也可以估计人体微生物菌群多样性累计最大值。同样理论,通过适当取样调查,也可以估计多样性随海拔高度变化累计最大值(例如喜马拉雅山从山基到山顶累计多样性),或者海洋不同深度生物多样性累计最大值。
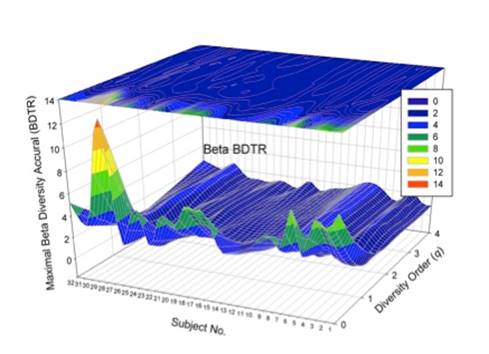
其三、类似于将SAR扩展为DAR,将STR(Species Time Relationship)扩展至一般的“多样性—时间关系”(DTR:Diversity Rime Relationship), 以及多样性时空关系(Diversity-Time-Area-Relationship)。它们也具有类似于DAR的应用如:监测(预测)人体微生物菌群动态变化。在应用于人体菌群时,可以为每个个体建立个性化DAR/DTR模型,从而对研究菌群相关疾病(例如肥胖)的个性化精准诊断治疗提供定量分析工具(如附图所示)。
其四、新提出的DAR/DTR/DTAR理论和模型不仅适用于Alpha-多样性、也适用于Beta-多样性、甚至Gamma-多样性,并且采用了统一的数学模型。
其五、所有提出的新概念和度量指标包括 DAR Profile, PDO Profile, MAD Profile 也均适用于Beta-多样性对时空尺度变化幅度的描述。所谓 Profile (系列特征)指的是:我们描述“多样性时空关系”(DAR/DTR/DTAR)其实是一个系列Renyi 熵值, 而不是像经典SAR/STR/STAR理论就是单纯一个物种数量。而系列内部多样性指数(也就是“希尔数”)之间的差别是由研究者对于物种丰度(Species Abundance) 看重程度(也就是“权重”程度)不同所决定。例如,如果研究或应用目的是珍稀动植物保护,则对像熊猫这样的稀有物种(Rare Species)的数量会增加权重(所谓高阶多样性、高阶熵、或者高阶希尔数)。而如果研究或应用目的是不同小龙虾品种的养殖,则权重自然应该倾向于普通丰度、甚至高丰度物种(Common Species)。这些新概念、模型和估计方法可以有效解决Southwood 所描述的、多样性评估中顾此失彼的困境。从而为研究多样性随时空尺度变化幅度的评估提供了灵活通用、功能强大的定量分析方法和工具。
团队采用了大量来自“人类微生物群系项目”(HMP, Human Microbiome Project)的微生物菌群数据,对于以上理论做出了广泛的验证。审稿专家对这一系列研究做出了高度肯定的评价。例如其中之一写道:“Technical quality of the work is high, and the paper is excellently written. I suggest expanding the description of the methods used for reconstructing the metagenomic datasets. In particular, it is important to specify the assembly strategy that was used for the computational demonstration” 。另外一位评价为:“Thus, this paper represents a pioneering paper that introduces some recently-developed ecological methodologies (especially the four classes Hill-number-based similarity measures) to metagenomic diversity analysis.” 编辑评论为:“This work is timely and important, and I have no problem accepting it for publication - well done on your research efforts. I've been telling my research group for years to explore the use of Hill's numbers, and I'm glad to see others providing a robust exploration of their uses when applied to metagenomic data.”
关于这些理论扩展和实际验证的具体工作,除[3]目前处于“修改”阶段外,其它将于近期发表在下列国际期刊, 并有四项国家发明专利进入实质审核阶段。对于重要计算过程,提供了相应的软件支持。该系列研究受到国家自然科学基金、云岭产业技术领军人才项目、国际合作项目—“基因调控变异预测及人类菌群宏基因大数据技术研发”、以及“遗传资源与进化国家重点实验室”等资助。
[1] Extending species-area relationships (SAR) to diversity-area relationships (DAR), Ecology and Evolution. (2018) https://arxiv.org/abs/1711.06231v1
[2] Diversity time-period and diversity-time-area relationships exemplified by the human microbiome, Scientific Reports. (2018) https://www.nature.com/articles/s41598-018-24881-3
[3] Sketching the Human Microbiome Biogeography with DAR (Diversity Area Relationship) Profiles, Microbial Ecology (in revision).
[4] Assessing and interpreting the within-body biogeography of human microbiome diversity; Frontiers in Microbiology. (2018) https://www.frontiersin.org/articles/10.3389/fmicb.2018.01619/abstract
[5] Measuring microbiome diversity and similarity with Hill Numbers. Chapter 8, in “Metagenomics Perspectives, Methods, and Applications” Academic Press. (2017)
https://www.sciencedirect.com/science/article/pii/B9780081022689000082
[6] Measuring metagenome diversity and similarity with Hill numbers. Molecular Ecology Resources. (2018) https://onlinelibrary.wiley.com/doi/10.1111/1755-0998.12923